
Did you know?
The largest hadrosaurid species discovered to date is Shantungosaurus giganteus. First described in China in 1973, it is estimated to have been forty-nine feet long and weighed seventeen tonnes. Its skull measured over five feet in length.
My research focuses on dinosaur evolution, feeding mechanics, and Mesozoic ecosystems.
During my Master’s, I chose to focus my thesis on hadrosaurid jaw evolution. These dinosaurs have always fascinated me, so I set out to examine their dentaries in detail to understand how their jaws changed over time and what evolutionary refinements shaped their success. The project proved insightful, engaging, and genuinely enjoyable, giving me a deeper appreciation for just how well adapted hadrosaurids were.
MSc Hadrosaur Mandibular Disparity Study
This study analysed 74 hadrosaur taxa to understand how jaw shape evolved across the Late Cretaceous.
Supervisors: Professor Mike Benton, Professor Emily Rayfield, Dr. James L. King.
Research Aims
My MSc thesis project focused on hadrosaur mandibular development across the Late Cretaceous, incorporating an Ornithopod outgroup. The project aimed to evaluate the role of feeding adaptation throughout hadrosaur evolution. I conducted a multivariate study of 74 hadrosaur taxa to track how dentary morphology evolved between 88–66 Ma.

MSc thesis. 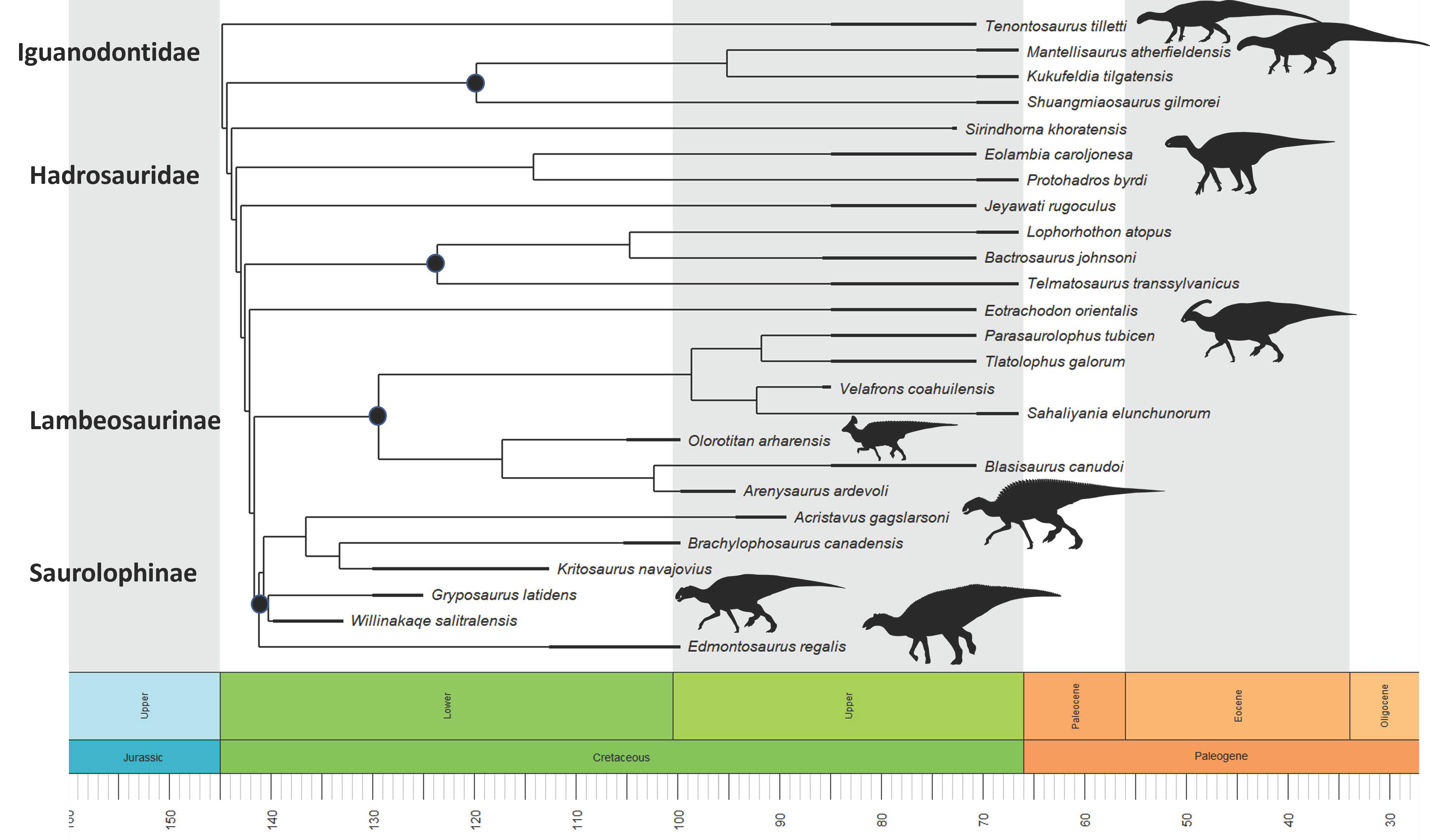
Ornithopod phylogenetic cladogram displaying 25 viable hadrosaur specimens of the study through geological time back to the Iguanodontids (Figure 1). 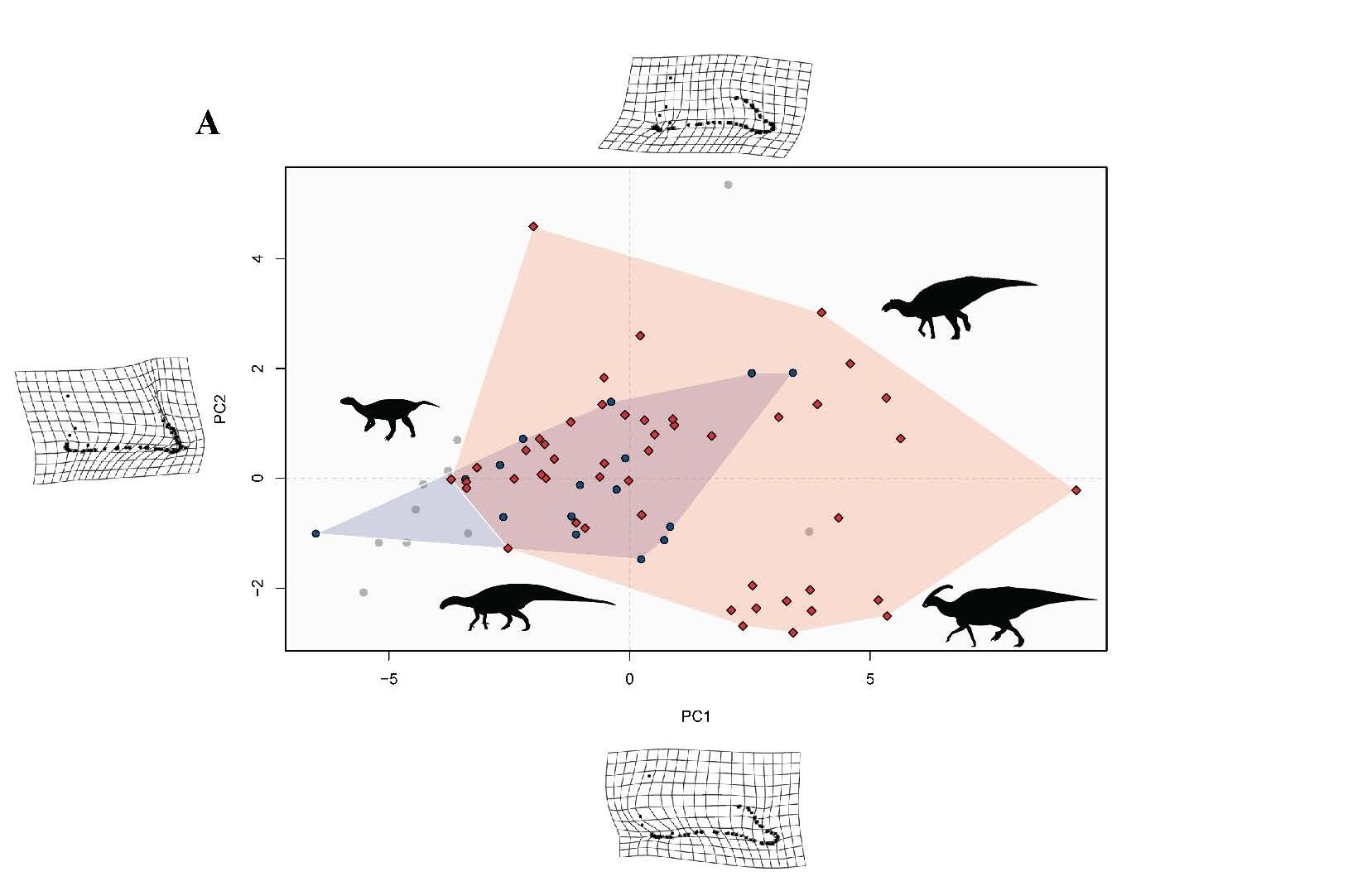
Principal component analysis (PCA) with 2D geometric morphometrics of the lower jaws of 74 hadrosaur taxa aligning to PC1 and PC2 (Figure 4). 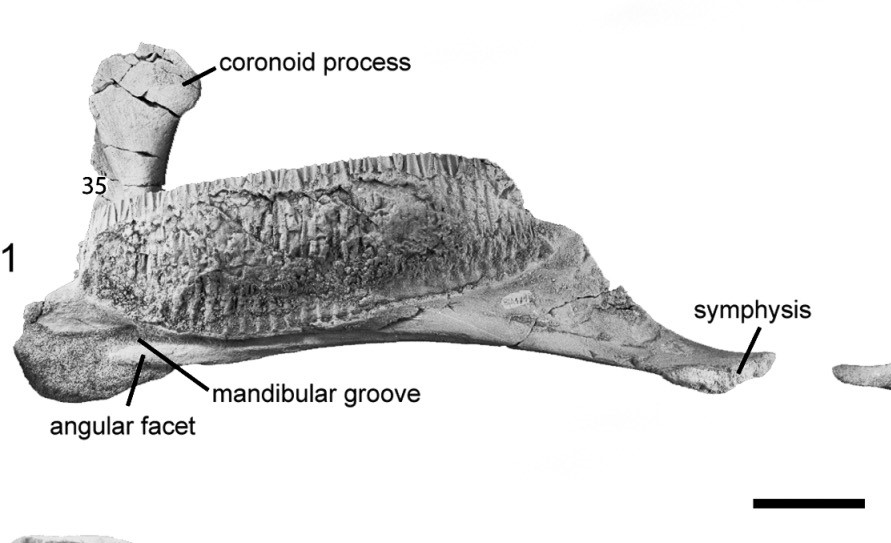
One of the lower dentary’s measured in the project from the lambeosaurine hadrosaurid Blasisaurus canudoi (MPZ 99/665). Scale bar is 5cm. Image credit: Caballero et al. 2010. 
Edmontosaurus annectens cast at Oxford Museum of Natural History. Edmontosaurus was one of the species used in the thesis project.
Methods
The study involved creating multivariate analyses of data in R studio, plotting a time calibrated phylogeny and morphospace plots displaying PC1 and PC2 jaw function systems for 74 hadrosaur taxa.
2D landmarking data was obtained using tpsDig, tpsRelw32 and tpsUtil32 for 29 hadrosaur specimens in the study. Assessing jaw shape change, with Generalised Procrustes Analysis (GPA) and Principal Components Analysis (PCA) normalising the shape data.
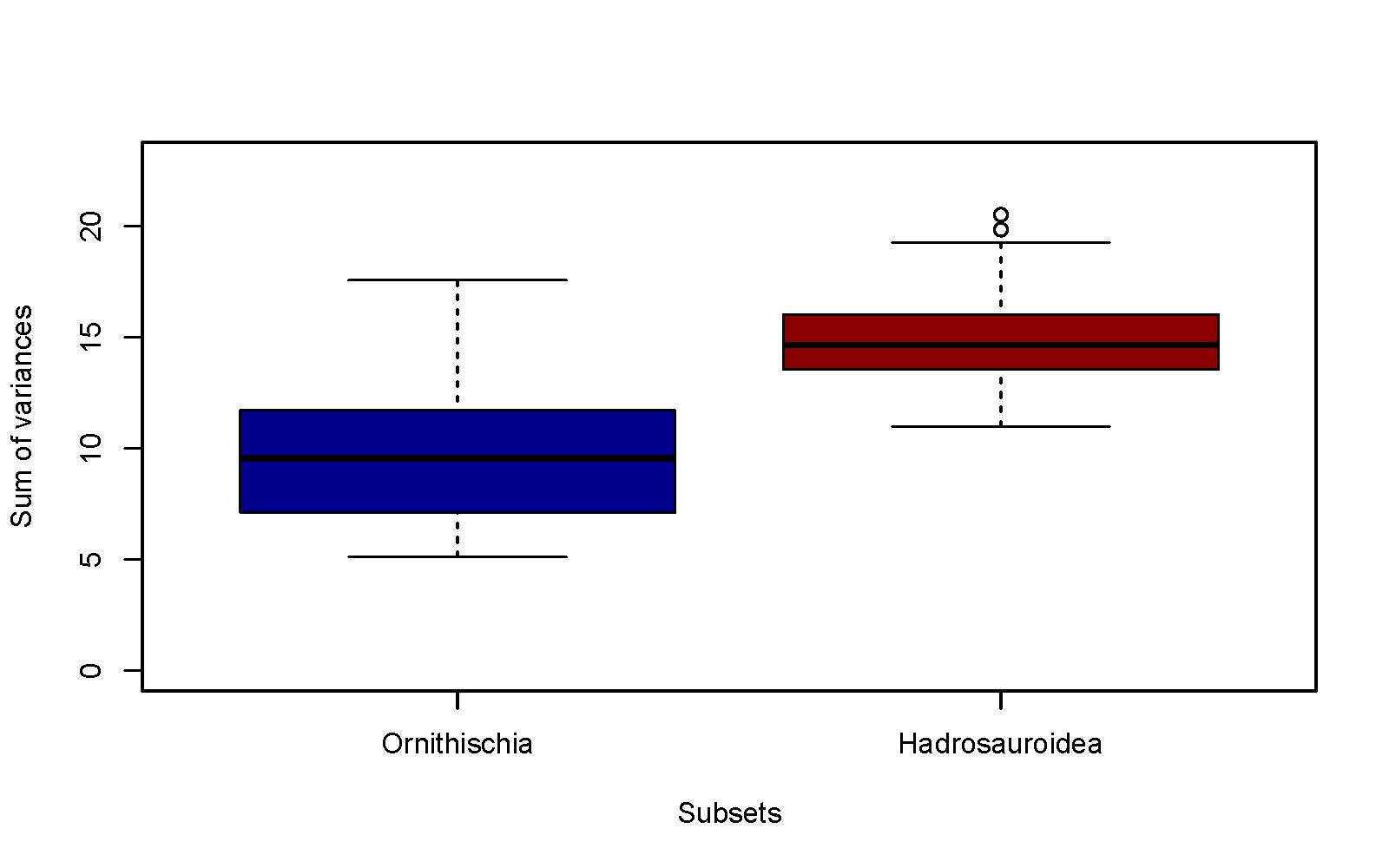
Histogram bar and box plots displayed jaw disparity within the clades. Timeline and time box plots were used to display times of significant variation of specimens across geological time.

Key Findings
Results suggested macroevolution of hadrosaurs on a grand scale with jaw specialisation found at the root of the non-hadrosaurid ornithischian clade. Variations of the angular facet, surangular and coronoid process suggested hadrosaurs developed more refined feeding apparatus across their evolution.
These developments led to specialisation and species richness across the Late Cretaceous enabling global dominance. The refinement of the dentary aided their success and helped hadrosaurs exploit their own ecological niches.
Details of the non-hadrosaurid ornithischian, hadrosaurid, and newly recognised ceratopsian specimens analysed in this study are displayed below (Table S1).
Specimen Dataset (Table S1)
Expand to view the complete specimen dataset used in this study.
You can click on the highlighted specimens below which will take you to their designated profile.
| Taxon | Family | FAD | LAD | Specimen |
| Dryosaurus altus | Dryosauridae | 150.8 | 145.5 | CM 3392 |
| Mantellisaurus atherfieldensis 1 | Hadrosauroidea | 130 | 125 | OUMNH PAL-K.59014b/p |
| Mantellisaurus atherfieldensis 2 | Hadrosauroidea | 130 | 125 | NHMUK R11521 |
| Bolong yixianensis | Hadrosauroidea | 125.45 | 122.46 | YHZ-001 |
| Jinzhousaurus yangi | Hadrosauroidea | 125.45 | 122.46 | IVPP V12691 |
| Sirindhorna khoratensis | Hadrosauroidea | 130 | 112.6 | NRRU3001-167 |
| Xuwulong yueluni | Hadrosauroidea | 125.45 | 112.6 | GSGM-F00001 |
| Eolambia caroljonesa 1 | Hadrosauroidea | 105.3 | 99.7 | CEUM 34357 |
| Eolambia caroljonesa 2 | Hadrosauroidea | 105.3 | 99.7 | CEUM 9758 |
| Jeyawati rugoculus | Hadrosauroidea | 94.3 | 89.3 | MSM P4166 |
| Eotrachodon orientalis | Hadrosauroidea | 85.8 | 84.9 | MSC 7949 |
| Acristavus gagslarsoni | Hadrosauroidea | 84.9 | 70.6 | MOR 1155 |
| Brachylophosaurus canadensis | Hadrosauroidea | 84.9 | 70.6 | MOR 1071 |
| Kritosaurus navajovius 1 | Hadrosauroidea | 84.9 | 70.6 | AMNH 5799 |
| Kritosaurus navajovius 2 | Hadrosauroidea | 84.9 | 70.6 | USNM 8629 |
| Secernosaurus koerneri | Hadrosauroidea | 83 | 66 | MACN RN 142 |
| Willinakaqe salitralensis 1 | Hadrosauroidea | 84.9 | 66.43 | MPCA-Pv SM 6 |
| Willinakaqe salitralensis 2 | Hadrosauroidea | 84.9 | 66.43 | MPCA-Pv SM 3 |
| Gryposaurus latidens | Hadrosauroidea | 85.8 | 70.6 | AMNH FARB 5465 |
| Gryposaurus monumentensis | Hadrosauroidea | 84.9 | 70.6 | RAM 6797 |
| Gryposaurus notabilis | Hadrosauroidea | 84.9 | 70.6 | MSNM V345 |
| Prosaurolophus maximus | Hadrosauroidea | 84.9 | 70.6 | CMN 2870 |
| Saurolophus angustriostris 1 | Hadrosauroidea | 70.6 | 66.43 | MPC 100/706 |
| Saurolophus angustriostris 2 | Hadrosauroidea | 70.6 | 66.43 | PIN 551/407 |
| Kundurosaurus nagornyi | Hadrosauroidea | 70.6 | 66.43 | AENM 2/846 |
| Edmontosaurus regalis | Hadrosauroidea | 70.6 | 66.43 | CMN 2289 |
| Edmontosaurus annectens | Hadrosauroidea | 70.6 | 66.43 | DMNH EPI1493 |
| Edmontosaurus sp | Hadrosauroidea | 70.6 | 66.43 | CM 38321 |
| Pararhabdodon isonensis | Hadrosauroidea | 70.6 | 66.43 | IPS SRA 27 |
| Tsintaosaurus spinorhinus | Hadrosauroidea | 84.9 | 70.6 | IVPP V725 |
| Parasaurolophus cyrtocristatus | Hadrosauroidea | 84.9 | 70.6 | DMNH EPV.132300 |
| Parasaurolophus tubicen | Hadrosauroidea | 84.9 | 70.6 | NMMNH P-25100 |
| Parasaurolophus walkeri | Hadrosauroidea | 84.9 | 70.6 | ROM 768 |
| Corythosaurus casuarius 1 | Hadrosauroidea | 84.9 | 70.6 | AMNH 5240 |
| Corythosaurus casuarius 2 | Hadrosauroidea | 84.9 | 70.6 | CM 11376 |
| Corythosaurus excavutus | Hadrosauroidea | 84.9 | 70.6 | TMP 1992.036.0250 |
| Hypacrosaurus stebingeri | Hadrosauroidea | 84.9 | 70.6 | MOR 549 |
| Magnapaulia laticaudus | Hadrosauroidea | 84.9 | 70.6 | LACM 20874 |
| Sahaliyania elunchunorum | Hadrosauroidea | 70.6 | 66.43 | GMH W451 |
| Velafrons coahuilensis | Hadrosauroidea | 84.9 | 70.6 | CPC 59 |
| Tlatolophus galorum | Hadrosauroidea | 73 | 72.5 | CIC/P/147 |
| Olorotitan arharensis | Hadrosauroidea | 70.6 | 66.43 | AEHM 2/845 |
| Hypacrosaurus altispinus | Hadrosauroidea | 70.6 | 66.43 | CMN 8501 |
| Arenysaurus ardevoli | Hadrosauroidea | 70.6 | 66.43 | MPZ2008-258 |
| Blasisaurus canudoi | Hadrosauroidea | 70.6 | 66.43 | MPZ 99/665 |
| Yamatosaurus izanagii | Hadrosauroidea | 71.94 | 71.69 | MNHAH D1-033516 |
| Lambeosaurus clavinitialis | Hadrosauroidea | 84.9 | 70.6 | VP.003222 |
| Lambeosaurus magnicristatus | Hadrosauroidea | 84.9 | 70.6 | CMN 8705 |
| Lambeosaurus lambei | Hadrosauroidea | 84.9 | 70.6 | CMN 2869 |
| Charonosaurus jiayinensis | Hadrosauroidea | 70.6 | 66.043 | CUST J-V1251-57 reconstruction |
| Iguanodon bernissartensis | Iguanodontidae | 145.5 | 112.46 | PAL-KZ.26569/p |
| Altirhinus kurzanovi | Iguanodontidae | 109 | 99.7 | PIN 3386/8 |
| Hypsilophodon foxii | Neornithiscia | 130 | 125.45 | BMNH R2477 |
| Anabisetia saldivia | Ornithischia | 93.9 | 92 | MCF-PVPH-74 |
| Tenontosaurus tilletti | Ornithischia | 112.6 | 99.7 | OMNH 58340 |
| Lanzhousaurus magnedens | Ornithischia | 129.4 | 125 | GSLTZP01-001 |
| Kukufeldia tilgatensis | Ornithischia | 139.8 | 132.9 | NHMUK 28660 |
| Batyrosaurus rozhdestvensk | Ornithischia | 83.6 | 72.1 | AEHM 4/1 |
| Hypselospinus fittoni | Ornithischia | 140.2 | 136.4 | NHMUK R1831 |
| Equijubus normani | Ornithischia | 112.6 | 105.3 | IVPP V 12534 |
| Protohadros byrdi | Ornithischia | 99.7 | 94.3 | SMU 74582 |
| Tethyshadros insularis | Ornithischia | 84.9 | 66.43 | SC 57021 |
| Bactrosaurus johnsoni | Ornithischia | 84.9 | 70.6 | AMNH 6553 |
| Gilmoreosaurus mongoliensis | Ornithischia | 84.9 | 70.6 | AMNH FARB 30654 |
| Shuangmiaosaurus gilmorei | Ornithischia | 105 | 99.7 | LPM0166 |
| Telmatosaurus transsylvanicus | Ornithischia | 70.6 | 66.43 | BMNH R3386 |
| Lophorhothon atopus | Ornithischia | 84.9 | 70.6 | YPM VPPU 023508 |
| Levnesovia transoxiana | Ornithischia | 94.3 | 89.3 | ZIN PH 466/16 |
| Probactrosaurus gobiensis | Ornithischia | 130 | 99.7 | PIN 2232/1 reconstruction |
| Haya griva | Parksosauridae | 83.6 | 72.1 | IGM 100/2017 |
| Changchunsaurus parvus | Parksosauridae | 112.6 | 93.9 | JLUM L0403-j-Zn2 |
| Rhabdodon priscus | Rhabdodontidae | 70.6 | 66.43 | N/A: Paper Allain & Suberbiola, 2002. |
| Zalmoxes robustus 1 | Rhabdodontidae | 70 | 66 | NHMUK R4912 |
| Zalmoxes robustus 2 | Rhabdodontidae | 70 | 66 | BMNH R.3392 |
| Mochlodon vorosi | Rhabdodontidae | 85 | 80 | MTM V 2010.105.1 |
Why This Matters
Hadrosaurids were highly social dinosaurs, well adapted to the environments they inhabited. Understanding their evolutionary success means examining how they evolved, adapted, and ultimately flourished. One key aspect of this is the development of their jaws, which were specialised for processing the vegetation of their time. Studying these feeding structures offers valuable insight into their palaeobiology and ecological success. With new fossils, discoveries, and analytical techniques emerging, we are gaining an increasingly detailed picture of the lives of these remarkable dinosaurs.
Together, these findings help clarify how hadrosaur feeding strategies evolved across the Late Cretaceous.
Learn More About Hadrosaurids
Join my premium newsletter for exclusive deep‑dives into hadrosaurid evolution and palaeobiology.

Triassic Microfauna of Hapsford Bridge, Vallis Vale, Frome
Before beginning my Master’s, I took part in the At the Feet of the Dinosaurs Internship Programme. Designed for students interested in palaeontology, the programme helped develop key research skills through the analysis, identification, sorting, and description of fossils from a Mesozoic locality, work that ultimately contributed to a published scientific paper.
Unlike my later Master’s research which focused on the Late Cretaceous, this project focused on Late Triassic (Rhaetian) microvertebrate fossils. It was an engaging and rewarding experience that involved close collaboration with the University of Bristol and Bristol Museum. My project examined over 1,700 microvertebrate fossils from the Vallis Vale SSSI to reconstruct the Late Triassic ecosystem and understand patterns of faunal diversity and preservation.
Supervisors: Professor Mike Benton and Dr Chris Duffin.
Project Background
This was a project I worked on with Professor Mike Benton and co-authors as an Honorary Research Associate at the University of Bristol. Working within the Palaeobiology Research Group the project saw me take part in fieldwork, identifying, sorting, and describing the Rhaetian age microvertebrate fossils taken from limestone samples at Hapsford Bridge, Vallis Vale.
Geological Context
The geological site of Vallis Vale is a Site of Specific Scientific Interest (SSSI) displaying Mesozoic sediments which rest on Carboniferous limestone. The stratigraphy begins with the Rhaetian in the east end, stepping up to the Middle Jurassic at the west end of the Vallis.

Vallis Vale completed paper. Image credit: Ronan et al. 2020. 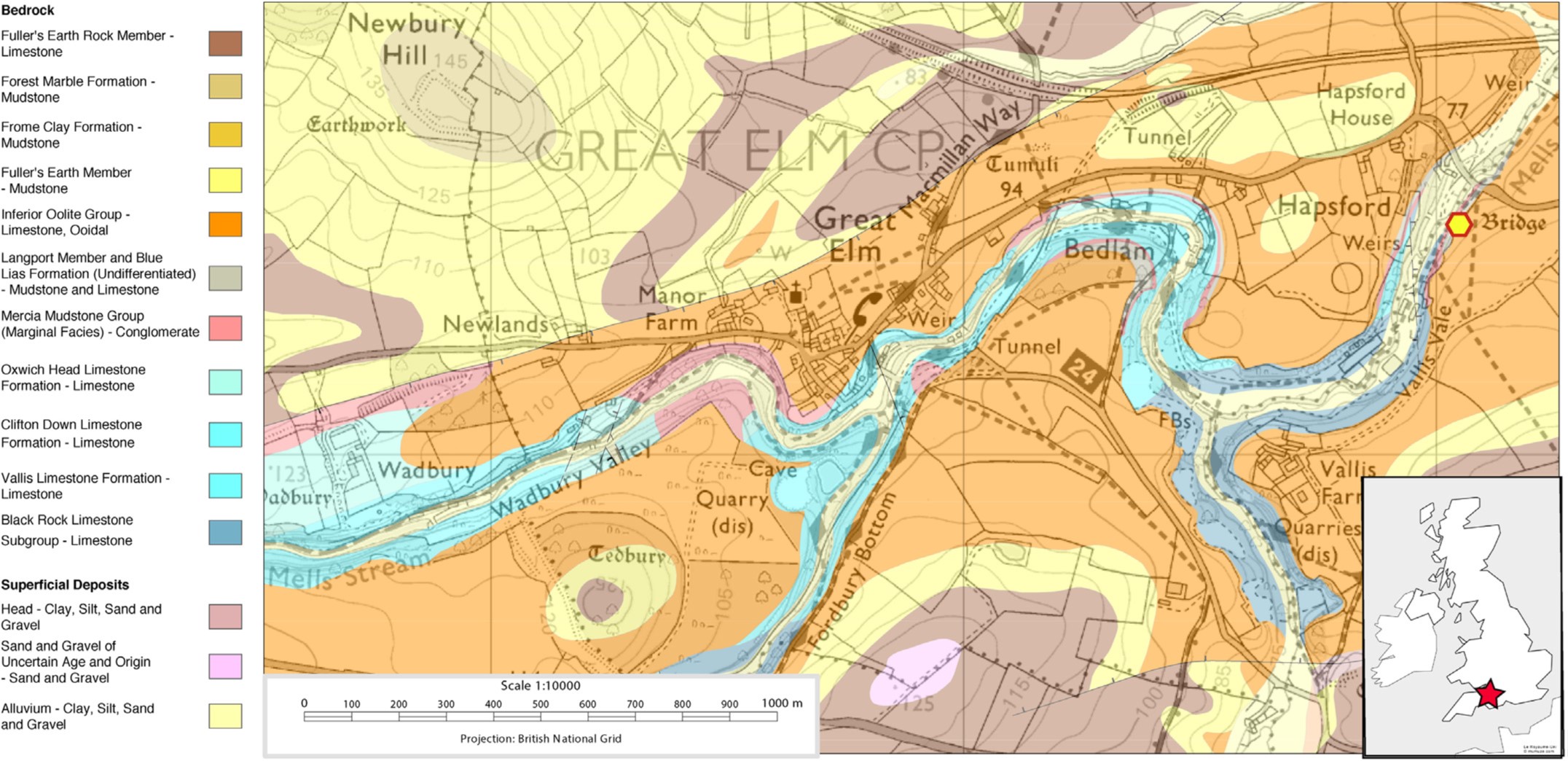
A geological map of Vallis Vale (Figure 3). Image credit: Ronan et al. 2020. 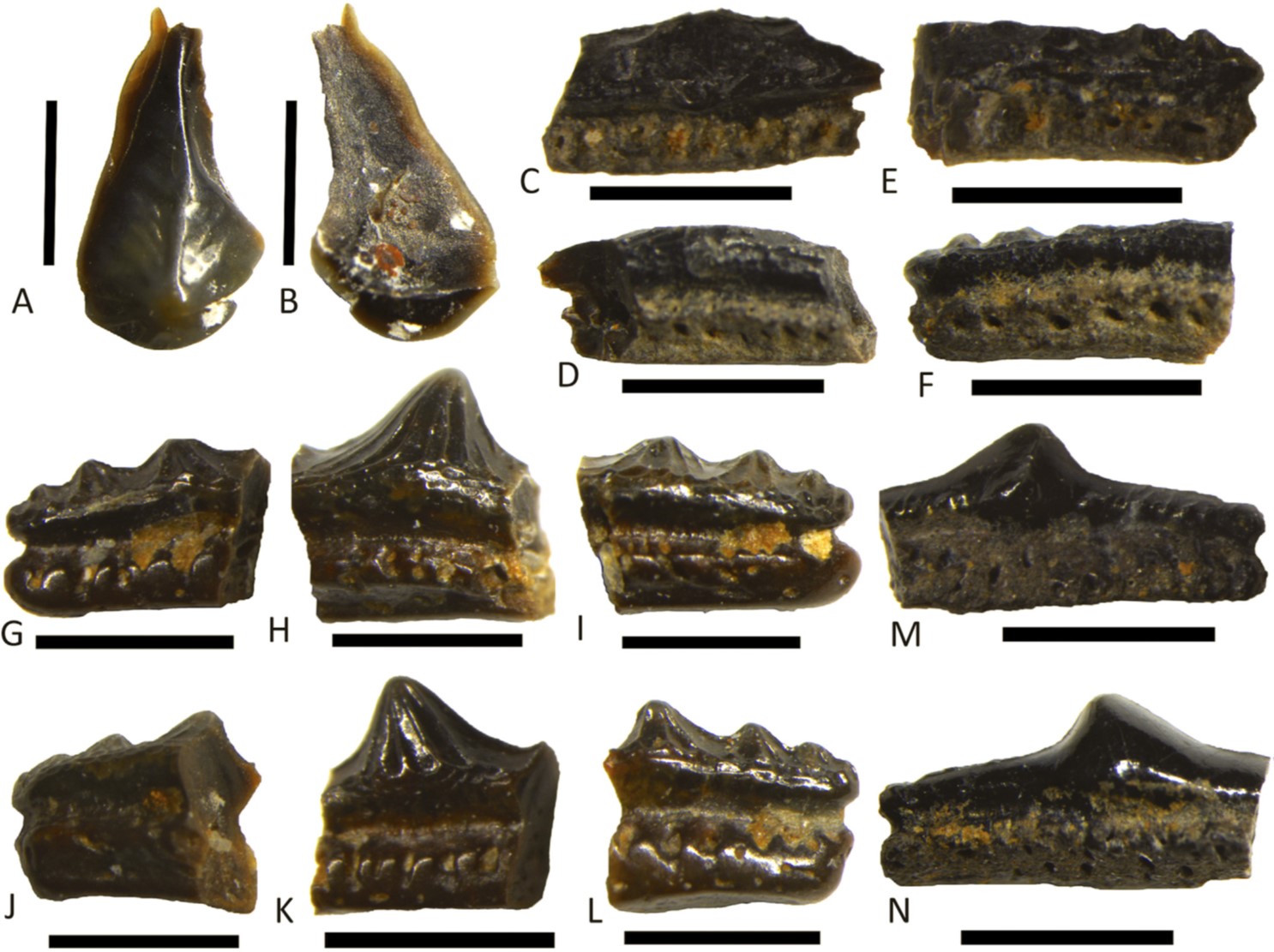
Chondrichthyan teeth identified in the study (Figure 6). Scale bar is 1mm for all specimens. Image credit: Ronan et al. 2020. 
Actinopterygian teeth and scales identified in the study (Figure 8). Scale bar 1mm for all specimens. Image credit: Ronan et al. 2020. 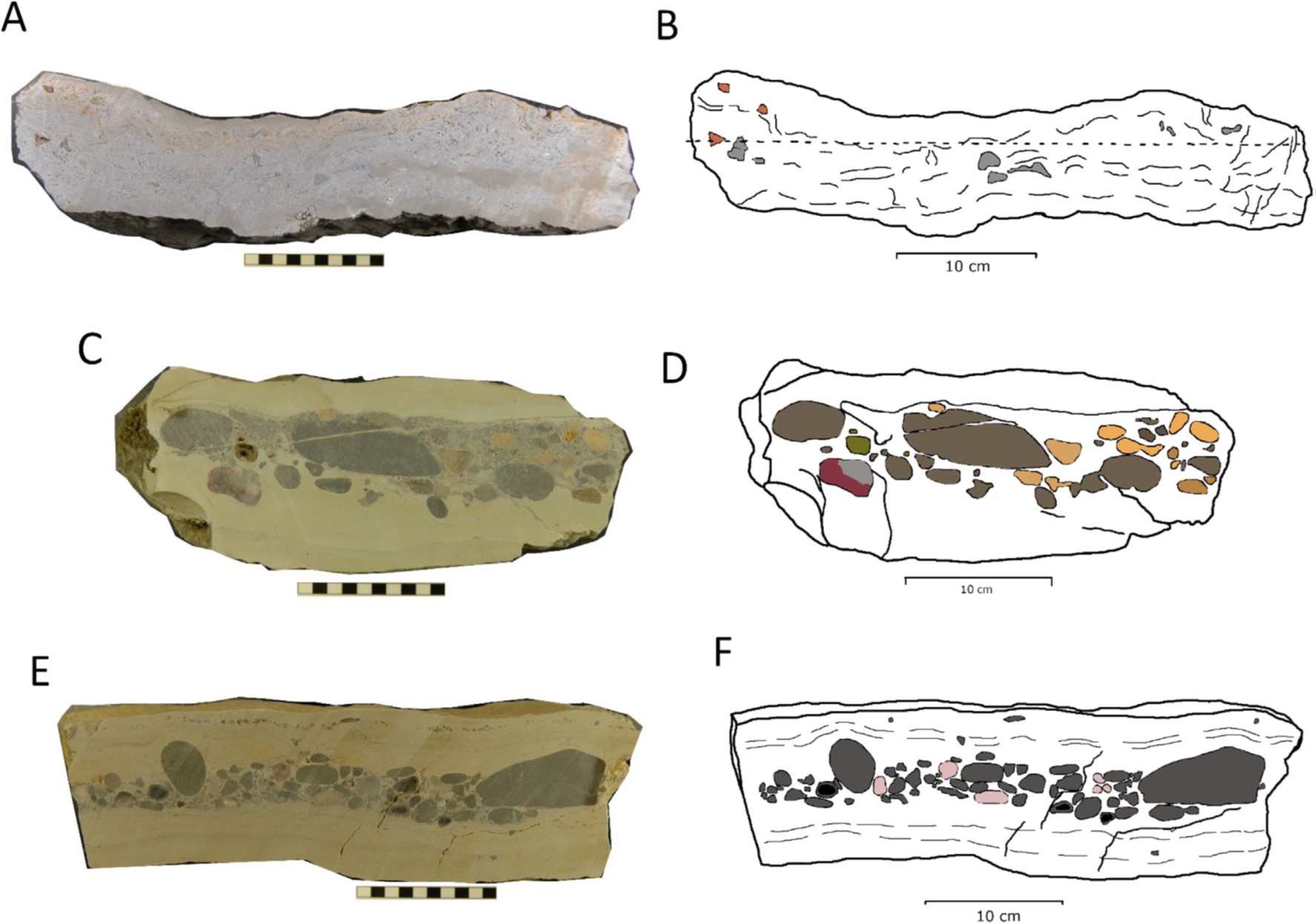
Inferred storm beds from different levels in the Westbury Formation (Figure 13). Image credit: Ronan et al. 2020. 
Presenting my Vallis Vale project research with Professor Mike Benton at Mendip Rocks! 2019.
Fossils Identified
The fossils collected were supported by fossil collections from Charles Copp, helping to display the Rhaetian Transgression which enveloped over the Carboniferous limestone at Vallis Vale.
The project identified 1,740 specimens with 763 non-bone fragments including 76 teeth and 687 other fossils elements. The Rhaetian bone bed layers display a variety of microverterbrates including four species of Triassic shark and two species of bony fish, alongside invertebrate fauna such as bivalves, echinoids, and trace fossils.
Results of the project can be accessed at the journal article here.
Why This Matters
The Vallis Vale research provides new insight into the palaeotopography and marine microvertebrate assemblages of the Mendips. The Rhaetian Transgression reshaped the landscape, and this study helps build a more detailed picture of the marine and geological changes that followed. Evidence of flooding, storm bed-deposition and climate shifts reveals how different marine faunas were mixed and preserved. The Triassic seas were teeming with life, and Vallis Vale offers a valuable snapshot of this dynamic ecosystem.
Together, these findings reveal the palaeoenvironmental conditions and the diverse marine microvertebrate fauna that inhabited the region 205 million years ago.
Explore More Dinosaur Work
Explore more of my dinosaur‑focused projects, hadrosaur insights, and my consultancy work.



Discover My Fieldwork Adventures!
Fieldwork is a core aspect of palaeontology. Explore how my fieldwork adventures have impacted research at the fieldwork adventures page.

Support the Science. Join the Expedition. Explore deep time with every dispatch. Subscribe to The Palaeo Minute newsletter and be part of the fossil journey.


You must be logged in to post a comment.